[古脊椎所]温血的起源
鸟类这一支的内恒温是什么时候出现的呢?由于所有现代鸟类都是内恒温,那么它们的最近共同祖先也可能是。bakker提出恐龙从祖先那里得到了温血,所有恐龙都是温血动物。而有人认为有些鸟类还不是内恒温,内恒温出现晚于鸟类飞行。还有人认为有些恐龙是内恒温,而在恐龙的主龙类祖先中有一定程度的内部调节体温的能力。如果内恒温不是所有恐龙具有的,那么应该问哪些恐龙可能是内恒温的。
长久以来恐龙被认为是冷血、迟缓的动物。1972年bakker主要基于恐龙与哺乳类及鸟类运动器官的相似性,以及捕食者与被捕食者的比率提出恐龙是内温动物。根据恐龙足迹可以得知恐龙是直立行走(即四肢位于身体下方而非侧方),他推测大型恐龙速度可以达到30千米/小时,而有些两足行走的可以达到50~80千米/小时。他还引用恐龙骨骼具有密集的哈氏管和层状骨来支持自己的结论。后来bakker将自己的观点扩充,在1986年出了一本科普读物《恐龙异端:新理论揭开恐龙的秘密和它们的灭绝》。他在其中扩充了证据,从多方面来论证,包括:恐龙成种和进化速度很高,生长速度快;温血代谢是顶级捕食者以及大型植食动物的进化优势,如果恐龙不是,就不会成为中生代的主角;一些恐龙居住在很靠北的地方,那里冷血动物很难暖和起来,所以恐龙应该是温血动物;大型恐龙如腕龙为了将血液输送到头部,心脏应该很高效;而温血动物的心脏比冷血动物的高效得多。
提出鸟类温血代谢的起源更可能发生在恐龙与主龙类间,而不是从恐龙到鸟类。他的学说提出后引发激烈争论,反对者甚众。
骨组织学研究是讨论脊椎动物温度调节机制的一个重要方法。曾经,恐龙的恒温骨组织学研究是基于外温动物与内温动物间骨组织学与生长速度以及基础代谢率存在相关性。如果一种动物生长方式类似现代爬行动物,缓慢生长,就被认为是冷血动物;如果迅速生长,类似哺乳类及鸟类,就认为可能是温血动物。bakker采纳的骨组织学证据被后来研究者认为是无效的。因为骨骼的高度血管化广泛分布在内温与外温动物中,哈氏管与热调节无关,而且纤维层状骨与高代谢可能无关。各类恐龙骨组织学特征差别很大,不能把全部恐龙同等看待。如有些恐龙在密质骨的外缘可见休眠线,而有的没有。这说明有些恐龙是有限定的生长,有的恐龙则是终生生长。基于很多恐龙中存在的生长环,chinsamy等人认为恐龙的代谢率介于爬行类与鸟类和哺乳类之间,热血的基本形式在鸟类的进化历史中出现得比较晚,并不是从它们的恐龙祖先那里继承下来的。而de ricqles和padian等认为低的生长速度并不能说明问题,恐龙,至少是兽脚类恐龙还是内温动物。他们的研究还推测翼龙有高的生长速度、高的代谢水平,有晚成雏,及育儿时间较长;也可能是内温动物。最近基于心脏的研究表明鳄鱼的祖先可能也具有一定的热调节能力,这样的话,鸟类的内温在基干主龙型类就已经初步具备了。
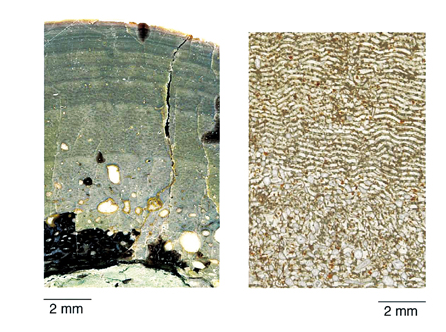
两种三叠纪恐龙的肱骨横切面
生物成因的磷灰石中氧同位素比率( 18op)很早就被用于重建古气候,但是在上世纪90年代研究者开始注意到其组成与脊椎动物体温有关,可以用于推测动物的代谢情况。起初有人根据骨骼中较均一的氧同位素比值推测恐龙是恒温的,但是有人质疑化石未能够保存原始的同位素比值。另外一种方法是比较恐龙与外温动物的氧同位素比率。假设共存的动物摄入的水的成分一致,但是氧同位素分馏与体温有关,所以磷灰石中氧同位素比率应该不同。在全球范围内生物体内的氧同位素比率主要取决于摄入的水,而水主要来源是雨水。雨水中氧同位素比率随纬度升高和气温下降而减小,故随纬度升高外温内温动物 18op值均减小。因为磷酸盐与水氧分馏随温度降低而增大,所以在大于纬度50 处,外温动物的 18op值比内温的高,而在低纬度外温动物的 18op值比内温的低。用这种方法对晚白垩世四类恐龙研究表明当时它们都具有高而恒定的体温,推测都是内温动物。这也是很间接的手段,无法得出直接的体温。此外由于体液以及生物磷灰石的 18o受当地雨水成分、湿度、生物食性、动物生理等因素的控制,以及后期石化作用的影响,结果有些不确定。最近兴起一种13c-18o团同位素(13c-18o clumped-isotope)温度测量法,依靠的是碳酸盐中13c和18o结合形成13c-18o团的倾向,这种倾向与总体同位素组成无关,而受温度的控制,故被用于重建古温度。这种方法在犀、象、鳄、鲨等现代生物中得到很好的验证;分析猛犸象的釉质得到很好的结果(38.4 1.8 c),但是受成岩作用影响的齿质结果远低于正常温度。后来用这种方法分析现生动物的蛋壳,结果也很令人满意。对保存良好的窃蛋龙的蛋分析得出31.9 2.9 c的温度,比同层产出的非生物成因的碳酸盐温度高6 c,但是比现生大多数内温动物体温低;而泰坦龙得到37.6 1.9 c的温度。
grady等人收集了化石和现代脊椎动物的个体生长数据,包括了恐龙的主要类群。他们发现生长率和代谢率关系基本符合理论预测。考虑大小和温度的影响,恐龙的代谢率处于典型的外温与内温间,与现生的中温最接近。基于生理学模型,小型外温的恐龙(小于100千克)生活在中高纬度体温远低于30 c,对它们的选择压力最强。如果它们没有体表的保温层,就不可能是内温动物。根据模型推测内温是在具保温功能的羽毛出现之后才演化出来的。
有人认为间接证据都是不可靠的,只有与内恒温功能有直接联系的结构才能作为证据。现代鸟类羽毛有两个功能:飞翔以及保暖。故羽毛的存在通常被当作是内温的可靠标志。尤其是早期原羽毛结构,如在中华龙鸟、中国鸟龙当中,并不具有飞翔功能,也不似性炫耀,最合理的解释是保温。目前在窃蛋龙中有孵蛋的行为,这表明其体温应该已经高于环境温度,而且这种小型恐龙应该是内温动物。不过有人否认这些,他们认为唯一可靠的标志是呼吸鼻甲骨的存在。呼吸鼻甲骨能够减少热和水的流失以及冷却大脑;在几乎所有现生鸟类和哺乳类中存在,除了鸬鹚、鹈鹕、鲸类等少数例外。即使在有真正羽毛的始祖鸟中,由于有前颌骨孔的存在,开口向副鼻窦(paranasal sinuses),所以鼻道窄,可能鼻甲骨不存在。所以他们认为内温是在鸟类飞翔之后产生的,为了加强长距离的飞翔能力而不是热调节。

几种有毛或者毛状衍生物的动物:(a)恐龙(中华龙鸟);(b)翼龙(宁城热河翼龙);(c)哺乳动物(玲珑仙兽)




