
关于作者
陈安国,浙江省临海县人。1958年7月毕业于北京大学生物系动物专业。先后在中国科学院动物研究所(北京)、西北高原生物研究所(西宁)和亚热带农业生态研究所(长沙)从事农业害鼠害虫生态与控制研究42年,主持完成中科院、省和国家的重大、重点、攻关、科学基金及国际合作项目的课题12项,参与8项。1992年10月起享受国务院“政府特殊津贴”。主笔或合作正式发表研究报告为主的科技论文70余篇、专著4本,并写作科普作品20多件。代表作有《小家鼠生态特性与预测》(1993,科学出版社)、《长江流域稻作区重要害鼠的生态学及控制对策》(1998,《农业重要害鼠的生态学及控制对策》第3章,海洋出版社)等。与郑智民、姜志宽合作主编《啮齿动物学》(第二版于2012年由上海交通大学出版社出版)
“走为上”计:主动迁移扩散—北疆小家鼠大暴发揭秘⑸
陈安国
2016年03月28日“走”,是北疆小家鼠擅长的生存对策
檀道济三十六计,“走为上”是讲战争中面对困局,宜主动出走、转移。动物界不少物种的生存对策也取此招。北疆小家鼠为避免种群密度增加导致居住空间拥挤与食物短缺,或面对生境条件改变,除了前述“生育自动调节”之外,更高超的对策也是“走”:种群密度高了,走——扩散;栖息地恶化,走——迁出;他生境优化,走——迁入!不仅大暴发时因食物耗尽而成群转移,平常年份随生境季节性变化也会适时迁移——严冬降临前迁入人房,天气转暖植被茂盛时迁向田野,并随着不同作物播种、生长、收割的变化,还会随机更换栖息地。这种特强的季节性迁移习性,正是北疆小家鼠面对强烈大陆性气候求生存发展之最上策,使该鼠种得以随时占据最有利的生活地段,能更有效地发挥超强生殖力,尽可能扩张种群,从而超越当地其他鼠类,成为最富暴发性的优势种。郁闭度,是北疆小家鼠在作物地间迁移的导因
天山北麓农区3月中下旬开春之后,苜蓿生长,冬麦返青,小家鼠即随之由人房等越冬地迁来。4月以前麦地较裸露,小家鼠数量尚很低;5月起麦苗生长迅速,鼠数量就相应上升,6月中旬达高峰,捕获率可为4月上旬的10~12倍。7月上旬小麦趋成熟,麦地的小家鼠数量却开始明显减少;与此同时,春播的胡麻、油菜等已长成而郁闭,其内小家鼠数量激增。8月小麦收割完毕,胡麻、油菜,以及芝麻、花生、谷子、糜子及豆类等低秆密生作物陆续进入黄熟期,这些地里的小家鼠亦达高峰。玉米、高粱和向日葵是夏播高秆作物,至8月长高郁闭,鼠数量始大幅上升;此后这类作物地少有人畜活动,下草茂盛,成为小家鼠增殖大本营,数量持续猛升,到10月收割前可达6、7月的20~40倍。当地水稻田多开垦于潜水溢出带,与旱作物地及居民点距离较远,分丘毗连,田埂纵横交织,多年不翻动,杂草茂盛,因而终年有小家鼠栖居。冬春稻茬地就有一定数量的小家鼠,而从7月水稻分蘖苗露出水面,到成熟收割的10(11)月,该鼠数量直线上升,通常高于其他作物地同期捕获率。
耕作区的另类生境是各种大小渠道、休闲地、撂荒地及草甸,总称“田间荒地”,杂草丛生,少受人干扰,但其籽实及植株养分品质较次,食物条件不及作物地。其上通常有灰仓鼠、沙鼠栖息,也会有一些小家鼠居住。特别是作物收割,这些田间荒地就成了小家鼠避难所与迁移中转站。
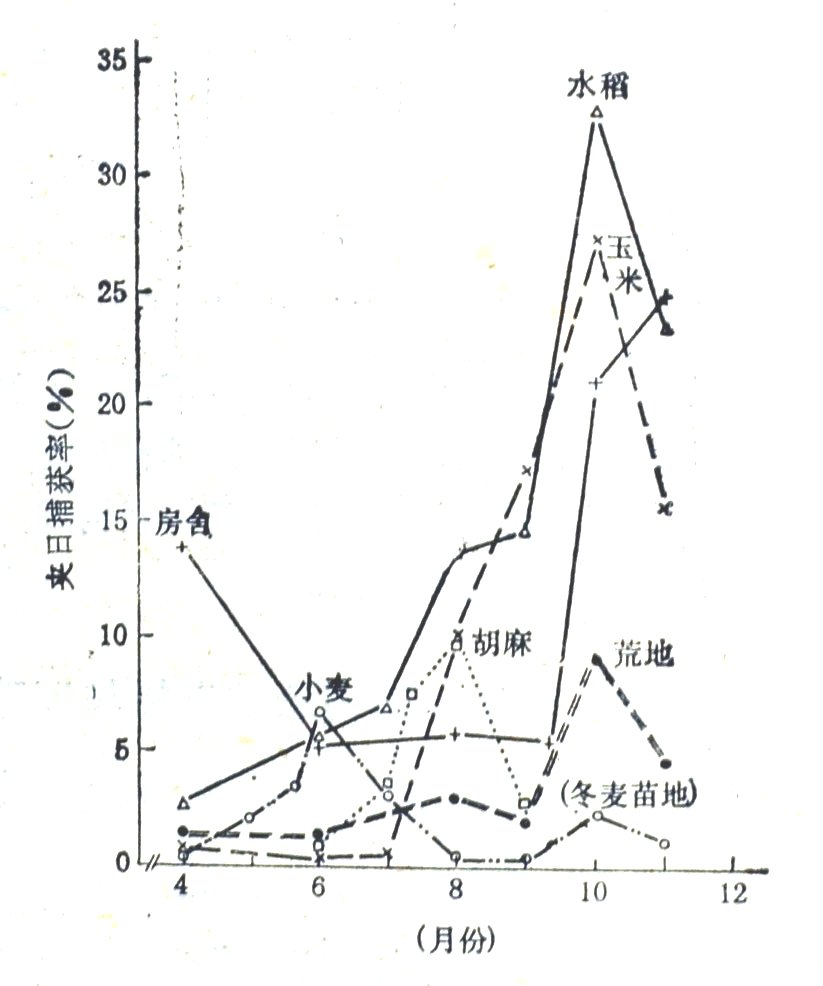
图5-1.6种代表性栖息地内小家鼠数量季节消长关系(玛纳斯,1970-1979)
图5-1是6种代表性栖息地内小家鼠数量季节消长关系图,除了房舍区曲线呈u形之外,4作物地的鼠数量都随迁入与繁殖而走高,并相继达高峰;6-8月的小麦地对胡麻地,8-9月的胡麻地对玉米地,曲线走向显著负相关,正是前地小家鼠向后地迁移的反映。玉米地与水稻田的小家鼠都是在秋末达顶峰,因收割而激降;随之,先荒地过渡,后进房舍区,这2个非作物地生境在秋末鼠数量激增。
应当注意的是,鼠类选择栖息地,一般讲是以丰富的食物、适宜的居住条件和良好的安全环境(少种间冲突)为目标,但北疆小家鼠在田野的迁移的直接导因则是植被郁闭度。
这可由多方面表现看出来。先看开春发端,北疆大地最先变绿的是苜蓿地和冬麦地,也就最先出现越冬地迁来的小家鼠(首批4月中旬捕获的都是成年鼠),非粮作物苜蓿地先长茂密,进鼠也就最多。再看麦地小家鼠动态,历年数量高峰都出现在刚全郁闭的灌浆期,到7月黄熟期反而锐减,这是因为此时胡麻等春播作物地也已郁闭,流动性强的小家鼠就扩散了,而流动性弱的灰仓鼠在此期间捕获率仍持续上升。新鲜麦粒的吸引力未能阻滞小家鼠向外扩散,恰好表明郁闭度对其作用更大。三看长势不同的低秆密生作物地,糜子为夏播作物,7月下旬尚处拔节期,未郁闭,小家鼠数量与胡麻地相差悬殊,到8月中旬抽穗期就无大差别了。而油菜地不同地块长势差别大,郁闭度高的,小家鼠数量亦高。四看玉米地,4月中下旬播种,经间苗和中耕除草,株、行距很大,至7月中下旬扬花时植株已高达1.5米,但未能郁闭,地内虽有些小家鼠并处繁殖盛期,历时3个月增长幅度一直很小;到8月,玉米长至2米且下草生长茂盛,形成良好的郁闭植被,小家鼠数量即猛增,但下草稀少地块鼠仍较少。这些事实一致表明,田野小家鼠的聚集与扩散,是以植被郁闭度作指向的。这应是由于各作物地的食物都会充裕,隐蔽条件就成了主要限制因素;何况郁闭度高,通常也意味着长势好、温湿度适宜,可利用的食物(包括草籽与虫类)及安全空间也相对较多。
房舍区,是北疆小家鼠严寒避难所
北疆冬季酷寒,石河子片上世纪50-70年代记录,1月平均气温为-16.8 ,日最低气温≤0.0 持续期161.6天,极端最低气温-39.8 。当地11月上旬入冬,小家鼠则先行随秋作物收割迁向碾场与房舍区,使人房内鼠密度在10-11月份激增。由于能继续繁殖,全冬鼠数量居高不下。但开春后它又主动迁出,5月鼠数量剧降,季节消长曲线呈u形。.png)
图5-2. 房舍内小家鼠数量季节消长曲线(玛纳斯)
这种主动外迁表明,北疆小家鼠仅将人房作为越冬避难所,农田作物长成它仍要回归。这乃是房舍內受人类防治压力大,作物地则活动空间和食物营养条件更佳。田野捕获的小家鼠,年平均胎仔数7.86只(8月份达8.56只),而房舍区捕获的,年平均胎仔数仅6.19只(8月份7.70只),比田野低21.2﹪。这既有气候因素(人房样本以冬季捕获的偏多),又有营养因素和栖息空间宽松的作用。所以,北疆小家鼠不恋“家”,它更喜走进宽阔的农田。反观内地如湖南的小家鼠乃偏安房舍,少有到野外栖息,因而生存空间较窄,难以形成暴发性数量。两相对照,足见北疆小家鼠善“走”确是生存竞争之上策,显具优越性。
图5-2乃显示,1972小暴发年,人房鼠数夏季即偏高,秋后更是异常高;1973为负反馈抑制年,全年数量皆低。1970年11月曾进行毒饵灭鼠,但几个月后即回升,表明单一化学灭鼠作用有限。
野外越冬,存活率极受气候制约
秋末作物收割后,仍会有小部分鼠留在秋作物茬地与田间荒地中。草茂、落穗落粒多的留居小家鼠也多,尤其是稻茬地、晚种晚收的糜子茬地及多杂草的老渠道,可以成为野外越冬地。厚厚的雪被对它们起保护作用,自11月至翌年1月,虽陆续有冻饿死的,仍会保有相当数量。玉米茬地、撂荒地起初鼠数亦多,因可藏身的田埂少、食物也少,一经放牧牛羊践踏,存量会迅速减少。野外越冬小家鼠大批量死亡发生在2月份,此时食物已很贫乏,鼠体质极度衰弱,气温却变化无常,时而暖至积雪消融,时而寒流突袭,能存活的就寥寥无几了。
分析上世纪70年代天山北麓农区每个冬季的气候,小家鼠大暴发的1967年、小暴发的1972与1974年,其前冬或积雪特厚或气温地温特暖,冬末春初气候平稳,加之上年的年成好,作物与杂草入冬时遗留籽实多。在这些有利条件保障下,野外越冬鼠存活率高;虽然鼠密度远不及室内,但由于面积大,汇聚到春作物地总数就很可观,致种群开春基数特高,奠定了那几年暴发的基础。
掌握迁移规律,才能准确监测和适时适地歼灭之
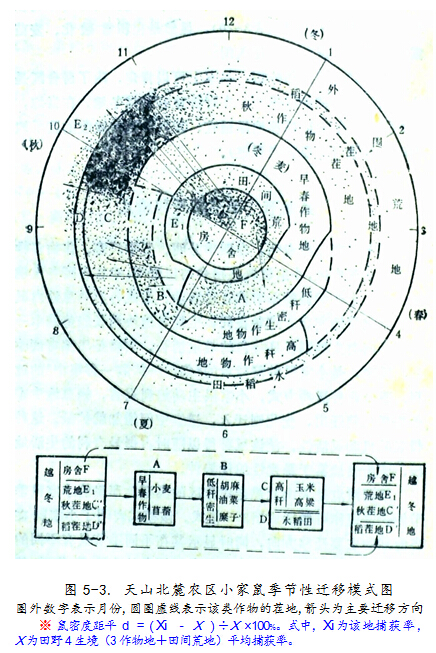
图5-3是依上世纪70年代调查资料描绘的天山北麓农区小家鼠季节性迁移模式。由该图可知,每个季节小家鼠种群都有一定的“密集地”,鼠密度距平※>10﹪以上。4月是稻茬地和田间荒地,6月是小麦地,8月是水稻田和胡麻地,10和11月是水稻田和玉米地(或两者的茬地)。其中,密度距平>50﹪的可视为该时期小家鼠“最适栖息地”,4月为稻茬地,6月为小麦地(及苜蓿地),10月为水稻田和玉米地,11月又是稻茬地。房舍区11月至翌年4月小家鼠捕获率都超出田野的平均密度1倍以上,显然是冬期小家鼠密集地和最适栖息地。因此,各时期鼠情监测重点就应瞄准这些生境。
鼠害的发生和发展是一环套一环的,一旦预测到小家鼠数量超过防治指标,应抓住它在某生境密集时予以聚歼。一般讲,6月中旬小家鼠大部集中在小麦地,同时稻茬地和房舍内还有较高密度的鼠,是人工灭鼠(特别是组织毒饵灭鼠)最好时机;此后它扩散了,总数量也增长了,就很难消灭。入冬后,小家鼠大量进入房舍区,宜在12月份组织聚歼,既可防止冬季室内鼠害,又可大大压低来年的种群增殖基数,收到事半功倍之效。
鼠害的发生和发展是一环套一环的,一旦预测到小家鼠数量超过防治指标,应抓住它在某生境密集时予以聚歼。一般讲,6月中旬小家鼠大部集中在小麦地,同时稻茬地和房舍内还有较高密度的鼠,是人工灭鼠(特别是组织毒饵灭鼠)最好时机;此后它扩散了,总数量也增长了,就很难消灭。入冬后,小家鼠大量进入房舍区,宜在12月份组织聚歼,既可防止冬季室内鼠害,又可大大压低来年的种群增殖基数,收到事半功倍之效。